Veronika Sander, Bruno Reversade and E. M. De Robertis
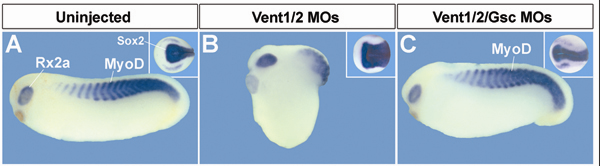
We present a loss-of-function study using antisense morpholino (MO) reagents for the organizer-specific gene Goosecoid (Gsc) and the ventral genes Vent1 and Vent2. Unlike in the mouse Gsc is required in Xenopus for mesodermal patterning during gastrulation, causing phenotypes ranging from reduction of head structures-including cyclopia and holoprosencephaly-to expansion of ventral tissues in MO-injected embryos. The overexpression effects of Gsc mRNA require the expression of the BMP antagonist Chordin, a downstream target of Gsc. Combined Vent1 and Vent2 MOs strongly dorsalized the embryo. Unexpectedly, simultaneous depletion of all three genes led to a rescue of almost normal development in a variety of embryological assays. Thus, the phenotypic effects of depleting Gsc or Vent1/2 are caused by the transcriptional upregulation of their opposing counterparts. A principal function of Gsc and Vent1/2 homeobox genes might be to mediate a self-adjusting mechanism that restores the basic body plan when deviations from the norm occur, rather than generating individual cell types. The results may shed light on the molecular mechanisms of genetic redundancy.
|
GSC is required for the dorsalization caused by Vent1/2 knockdown. |
|
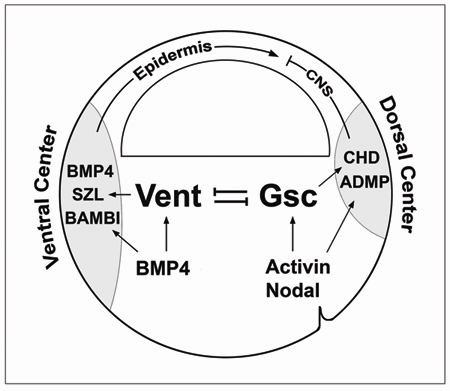
Model of regulatory mechanisms for pattern formation at gastrula. |
E. M. De Robertis
In: Gastrulation, C. Stern, Ed., Cold Spring Harbor Laboratory Press, New York, pp. 581-589 (2004)
For many years, the transplantation of Spemann's organizer into a host embryo was the only way of revealing the remarkable biological activity of the dorsal lip of the blastopore. The discovery of goosecoid (gsc) initiated the molecular era of research on Spemann's organizer (Cho et al., 1991). It became possible to visualize, by following gsc expression, the region of the embryo that corresponds to the inductive activity. The Spemann organizer became a concrete group of cells, rather than an embryological concept. Furthermore, since microinjection of goosecoid (gsc) synthetic mRNA is able to execute some of the properties of Spemann's organizer, these initial studies implicated homeobox genes as key elements in the function of the organizer. Subsequently many other homeobox genes were found to be expressed in Spemann's organizer. These transcription factors control the expression of secreted growth factors and growth factor antagonists that mediate the inductive activities of the organizer on neighboring cells. This chapter reviews studies on goosecoid and what they have taught us about the organizer, how embryonic signalling pathways are integrated at the level of the promoter, and the comparative anatomy of gastrulation.
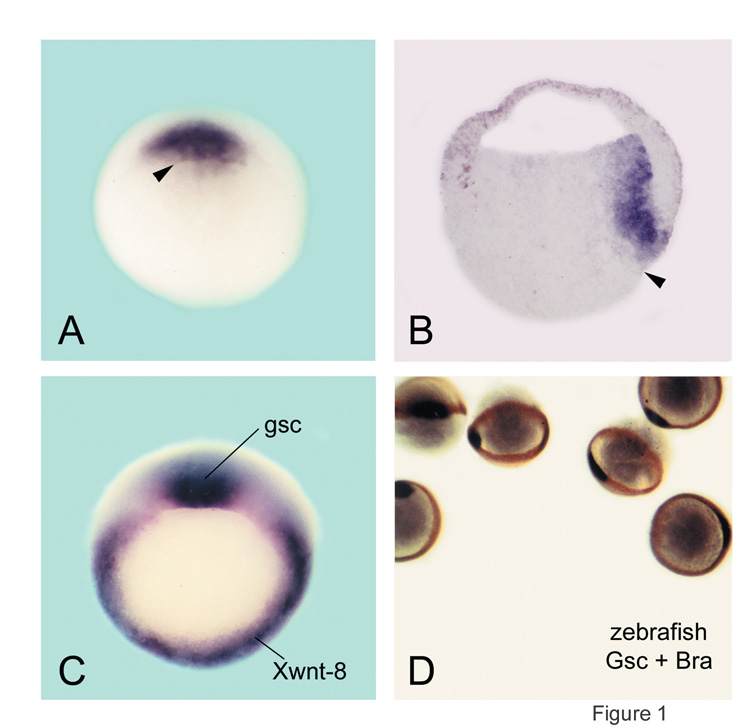 |
Fig. 1. Expression of goosecoid in Spemann's organizer in Xenopus and zebrafish. |
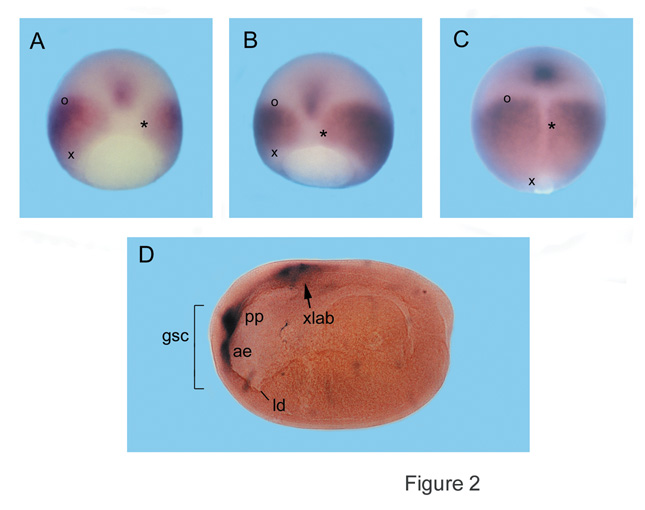 |
Fig. 2. Expression patterns of goosecoid and
the Xlab Hox gene during Xenopus gastrulation. |
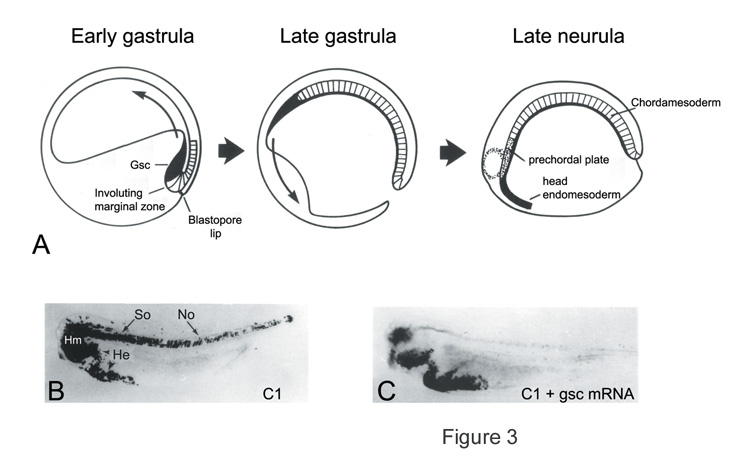 |
Fig. 3. Overexpression of goosecoid mRNA promotes dorso-anterior migratory movements. |
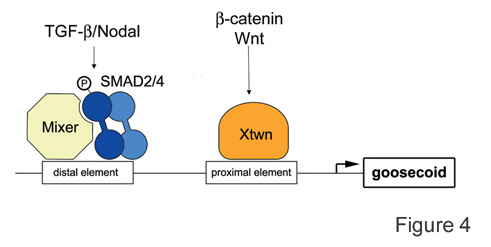 |
Fig. 4. Diagram of the goosecoid promoter region, showing how signals from the b-catenin and the TGF-b pathway are integrated at the level of the DNA. |
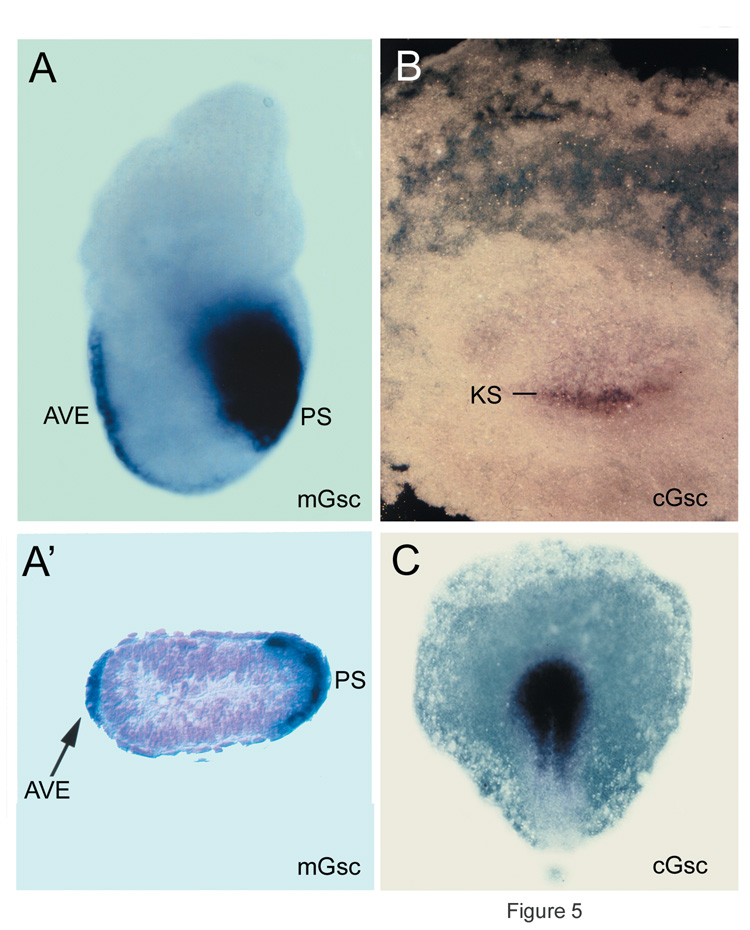 |
Fig. 5. Expression of gsc in mouse and chick embryos |
Ken W. Cho, Bruce Blumberg, Herbert Steinbeisser and E.M. De Robertis
This study analyzes the function of the homeobox gene goosecoid in Xenopus development. First, we find that goosecoid mRNA distribution closely mimics the expected localization of organizer tissue in normal embryos as well as in those treated with LiCl and UV light. Second, goosecoid mRNA accumulation is induced by activin, even in the absence of protein synthesis. It is not affected by bFGF and is repressed by retinoic acid. Lastly, microinjection of goosecoid mRNA into the ventral side of Xenopus embryos, where goosecoid is normally absent, leads to the formation of an additional complete body axis, including head structures and abundant notochordal tissue. The results suggest that the goosecoid homeodomain protein plays a central role in executing Spemann's organizer phenomenon.
Herbert Steinbeisser, Abraham Fainsod, Christof. Niehrs, Yoshiki Sasai, and E. M. De Robertis
Christof Niehrs, Herbert Steinbeisser and E. M. De Robertis
Herbert Steinbeisser, E. M. De Robertis, M. Ku, D.S. Kessler and Douglas A Melton
Development 118, 499-507 (1993)
Christof Niehrs, Rolf Keller, Ken W. Cho and E. M. De Robertis
Bruce Blumberg, Christopher V. Wright, E. M. De Robertis and Ken W. Cho
Martin Blum, S.J. Gaunt, Ken W. Cho, Herbert Steinbeisser, Bruce Blumberg, Dennis Bittner and E. M. De Robertis
S. J. Gaunt, Martin Blum and E. M. De Robertis
Development 117, 769-778 (1993)
G. Yamada, A. Mansouri, M. Torres, E.T. Stuart, Martin Blum, M. Schultz, E. M. De Robertis and Peter Gruss
Development 121, 2917-2922 (1995)
Jose A. Belo, Luc Leyns, G. Yamada and E. M. De Robertis
Mechanisms of Development 72, 15-26 (1998)
Gsc-1 expression marks cells with Spemann organizer, or axis-inducing, activity in the vertebrate gastrula. Gsc-1 knockouts, however, did not display phenotypes related to the early phase of expression. In this paper, additional phenotypes for the Gsc-1 mouse mutant are presented. Examination of the base of the cranium in the dorsal view revealed fusions and deletions in the midline of the prechordal chondrocranium. These defects were correlated with the sites of expression of Gsc-1 in the prechordal plate/foregut endoderm in the day 7.5/8.5 embryo. Gsc-1 expression in proximal limb buds was correlated with malformations of the shoulder and hip articulations. In addition, ribs in the seventh cervical vertebra were observed with low penetrance. The role of Gsc-1 during gastrulation and axial development is discussed in relation to possible compensatory interactions with other genes such as HNF-3beta and the recently identified Gsc-2 and Gsc-3 genes.
L. Zhu, Jose A. Belo, E. M. De Robertis and C. D. Stern
Developmental Biology 216, 276-281 (1999)
The homeobox gene goosecoid was the first specific genetic marker of Spemann's organizer in vertebrate embryos to be discovered. In the frog, misexpression of this gene by RNA injection produces duplication of the posterior axis. For these reasons, the recent finding that mice lacking goosecoid function have no early axial defects was rather surprising. Here we assay the neural inducing strength of wild-type and goosecoid-mutant mouse nodes by transplantation into primitive streak stage chick embryos. Wild-type mouse nodes strongly induce the neural-specific transcription factors Sox2 and Sox3 in the chick host. Homozygous goosecoid(-/- )nodes are severely impaired in their ability to induce both genes. Heterozygous goosecoid(+/-) nodes induce Sox3 as well as wild-type nodes, but resemble -/- nodes in their limited ability to induce Sox2. We propose that goosecoid does play a role in regulating the neural inducing strength of the node and that regulative mechanisms exist which mask the early phenotypic consequences of goosecoid mutations in the intact mouse embryo.
Stefan Schulte-Merker, M. Hammerschmidt, D. Beuchle, Ken W. Cho, E.M. De Robertis and C. Nusslein-Volhard
Development 120, 843-852 (1994)
Juan C. Izpisua-Belmonte, E.M. De Robertis, K.G. Storey and Claudio D. Stern
The chick homeobox gene goosecoid (gsc) is first expressed in a barely noticeable cell population near the posterior margin (Koller's sickle) of the unincubated egg. Then it is detected in Hensen's node, traditionally considered the chick organizer. Later, gsc-expressing cells leave the node with the prechordal plate. Fate mapping indicates that these three regions are related by cell lineage, and transplantation experiments suggest that they all have inducing activity. Quail posterior margin and anterior primitive streak grafts (gsc expressing) induce gsc transcription in neighboring chick host cells. We propose that development of the chick organizer starts earlier than previously thought and that gsc marks this changing cell population.
T.A. Heanue, R.L. Johnson, Juan C. Izpisua-Belmonte, Claudio D. Stern, E.M. De Robertis and Clifford J. Tabin
Mechanisms of Development 69, 31-37 (1997)
The homeobox-containing gene goosecoid (gsc) has been implicated in a variety
of embryonic processes from gastrulation to rib patterning. We have analyzed
the role it plays during chick limb development. Expression is initially observed
at stage 20 in a proximal-anterior-ventral domain of the early limb bud which
expands during subsequent stages. Later in limb development a second domain
of expression appears distally which resolves to regions which surround the
condensing cartilage. In order to understand the function of gsc in limb development,
we have examined the effect of misexpressing gsc throughout the limb. Two
striking phenotypes are observed. The first, evident at stage 24, is an alteration
in the angle of femur outgrowth from the main body axis. The second, which
can be detected at day 10 of development, is an overall decrease in the size
of the limb with bones that are small, misshapen and bent. These phenotypes
correlate with a decrease in levels of Hox gene expression in gsc-infected
limb buds. From these results we suggest that gsc may normally function to
regulate growth and patterning of the limb, perhaps through regulation of
Hox gene expression.

Last updated: March 2019