The MVBs and Wnt Page
______________________________________________________________________________________________________________________________________________
Wnt signaling requires sequestration of glycogen synthase kinase 3 inside multivesicular endosomes
Vincent F. Taelman, Radoslaw Dobrowolski, Jean-Louis Plouhinec, Luis C. Fuentealba, Peggy P. Vorwald, Iwona Gumper, David D. Sabatini and Edward M. De Robertis
Cell 143, 1136-1148 (2010)
Canonical Wnt signaling requires inhibition of Glycogen Synthase Kinase 3 (GSK3) activity, but the molecular mechanism by which this is achieved remains unclear. Here, we report that Wnt signaling triggers the sequestration of GSK3 from the cytosol into multivesicular bodies (MVBs), so that this enzyme becomes separated from its many cytosolic substrates. Endocytosed Wnt colocalized with GSK3 in acidic vesicles positive for endosomal markers. After Wnt addition, endogenous GSK3 activity decreased in the cytosol, and GSK3 became protected from protease treatment inside membrane-bounded organelles. Cryoimmunoelectron microscopy showed that these corresponded to MVBs. Two proteins essential for MVB formation, HRS/Vps27 and Vps4, were required for Wnt signaling. The sequestration of GSK3 extended the half-life of many other proteins in addition to β-Catenin, including an artificial Wnt-regulated reporter protein containing GSK3 phosphorylation sites. We conclude that multivesicular endosomes are essential components of the Wnt signal-transduction pathway.
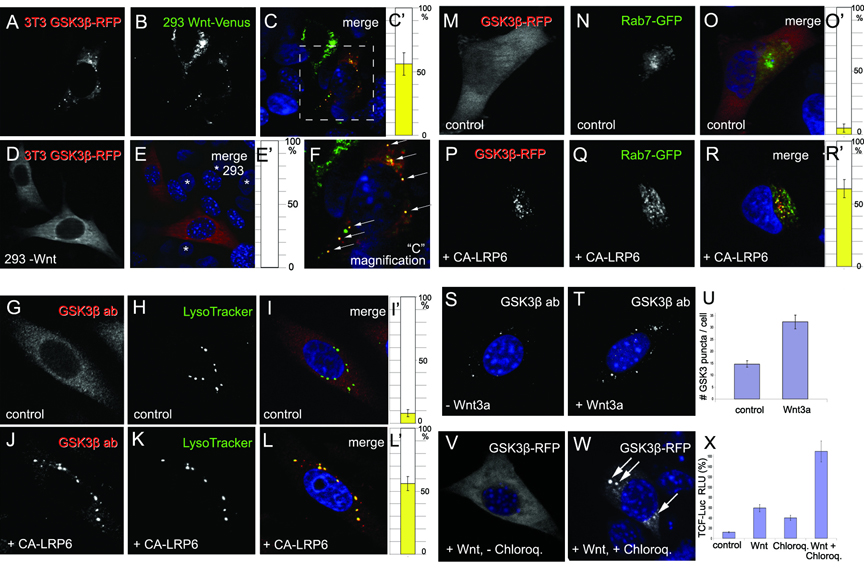 |
Figure 1. GSK3b is Translocated to Acidic Vesicles upon Wnt Signaling
(A-F) In co-culture experiments, xWnt8-Venus (green) secreted by 293T cells caused the relocalization of GSK3b-RFP (red) in 3T3 cells that endocytosed Wnt. Arrows in the enlargement in panel F indicate xWnt8-Venus endosomes that sequester GSK3-RFP. In controls lacking xWnt8-Venus, GSK3-RFP localization was cytoplasmic (D and E); asterisks indicate untransfected control 293 cell nuclei.
(G-L’) GSK3b antigen relocalized to LysoTracker-positive endosomes upon activation of Wnt pathway by CA-LRP6 in 3T3 cells; note that endogenous GSK3b was depleted from cytoplasm.
(M-R’) Rab7-GFP (green) and GSK3b-RFP (red) co-localized in CA-LRP6 signalosomes (in 62±7% of GSK3-positive puncta, n=80 HeLa cells); note GSK3 sequestration from cytosol.
(S-U) Endogenous GSK3b puncta (white) were larger and more numerous after 4 h of Wnt3a addition to untransfected 3T3-cells permeabilized with Digitonin. |
 |
Figure 2. Wnt Signaling Causes Sequestration of GSK3 inside Multivesicular Endosomes
(A) GSK3 kinase activity was decreased by 66±5% by Wnt3a treatment, and was recovered after membrane solubilization with 0.1% Triton X-100 in Digitonin-permeabilized L-cells. LiCl inhibition shows that the radioactive assay was specific for GSK3. Data from two independent experiments using untransfected L-cells. GSK3β and α-Tubulin provide loading controls.
(B) GSK3b becomes protected from Proteinase K after Wnt3a treatment, but only in the absence of Triton X-100 (lanes 3-6). Similar results were obtained in 5 independent experiments (untransfected L-cells). All samples were permeabilized with Digitonin, which causes leakage of 37% of the endogenous GSK3 (3 independent experiments).
(C and D) Cryoimmuno electron microscopy demonstrating relocation of endogenous GSK3β into MVBs (arrows) after Wnt3a treatment in untransfected 3T3 cells.
(E-G) GSK3-GFP localized in MVBs (white arrows) in CA-LRP6 transfected HeLa cells, but was cytosolic in control cells lacking CA-LRP6 transfection.
(H-J) Rab5-QL-DsRed forms giant endosomes (arrows), while GSK3β-GFP remains uniformly distributed in the cytosol (n=100).
(K-M) In the presence of CA-LRP6, GSK3-GFP is translocated inside Rab5-QL giant multivesicular endosomes (see arrows) in 77±9%, n=80, of co-transfected cells. Note that GSK3 becomes depleted from cytosol.
Data are represented as mean ± SEM.
|
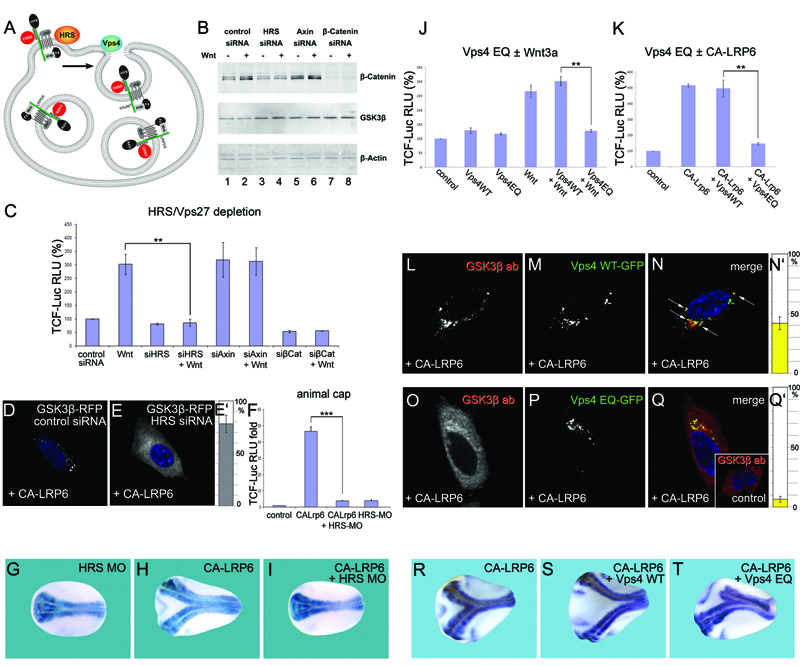 |
Figure 3. Components of the ESCRT Machinery Are Required for Wnt Signaling
(A) HRS and Vps4 are proteins required for intraluminal vesicle formation in MVBs. GSK3 is indicated in red.
(B) HRS siRNA inhibits Wnt3a-induced accumulation of β-Catenin.
(C) TCF-luciferase reporter gene assays showing that Wnt signaling requires HRS (brackets).
(D-E’) HRS is required for sequestration of GSK3-RFP in CA-LRP6 signalosomes (78±8%, n=150).
(F) Wnt signaling induced by CA-LRP6 mRNA in Xenopus animal caps was blocked by HRS morpholino (brackets).
(G-I) Induction of secondary axis in Xenopus embryos by CA-LRP6 mRNA (80 pg) requires HRS (neural tissue visualized with Sox2 probe).
(J and K) Expression of Vps4-EQ, but not Vps-WT, inhibited signaling by Wnt3a or CA-LRP6 (brackets) in 293T cells.
(L-N’) CA-LRP6 signalosomes sequestered endogenous GSK3β and partly co-localized with Vps4-WT-GFP, a multivesicular endosome marker (43±5% of vesicles, arrows, in n=80 cells).
(O-Q’) Overexpression of Vps4-EQ-GFP inhibits sequestration of endogenous GSK3b in CA-LRP6 signalosomes.
(R-T) Head formation in axes induced by CA-LRP6 was inhibited by Vps4-EQ, but not by Vps4-WT mRNA. |
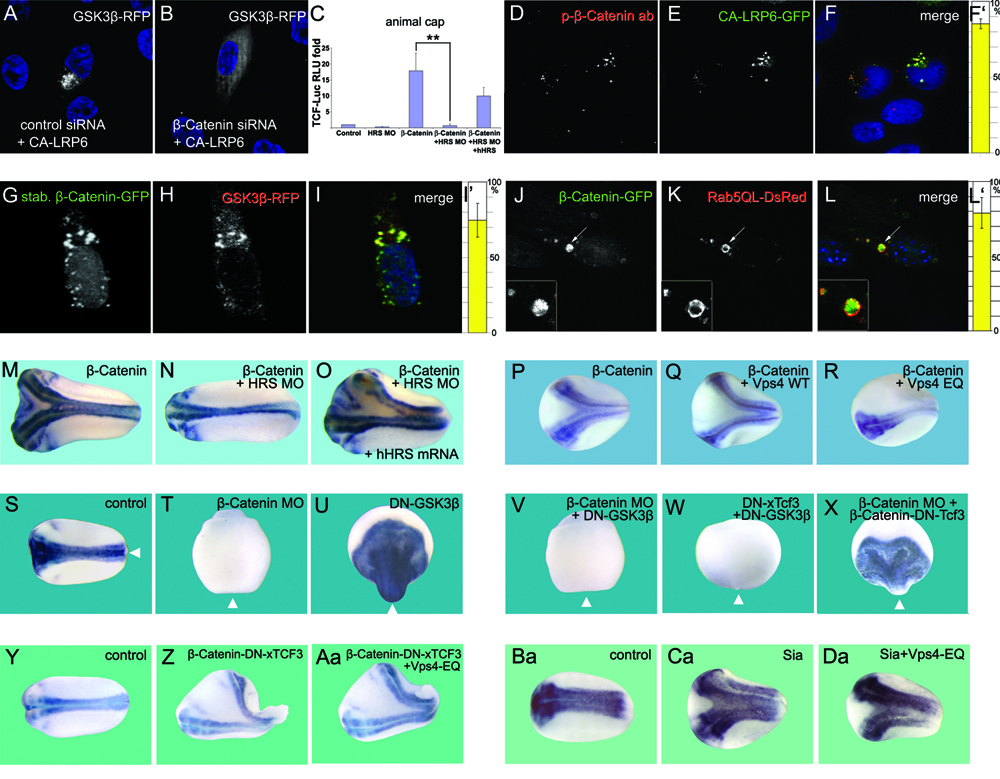 |
Figure 4. β-Catenin Is Required for GSK3 Localization in Multivesicular Endosomes
(A and B) β-Catenin siRNA inhibited GSK3β relocalization in CA-LRP6 signalosomes (in 86±7% of transfected HeLa cells, n=300).
(C) HRS MO blocks the induction of TCF reporter expression (brackets) by β-Catenin mRNA (4 injections of 80 pg) in Xenopus animal cap explants, and this was partially rescued by human HRS mRNA.
(D-F) Endogenous phospho-β-Catenin co-localizes with CA-LRP6-GFP signalosomes in HeLa cells (85±3% of transfected cells, n=56).
(G-I’) Overexpression of a stabilized mutant of β-Catenin-GFP caused its accumulation both in the nucleus and in cytoplasmic particles that sequester GSK3-RFP from the cytoplasm (75±11%, n=60).
(J-L) Wild-type β-Catenin-GFP becomes localized inside giant endosomes induced by Rab5-QL-DsRed. 79±10% of giant MVBs contained β-Catenin-GFP, n=80.
(M-O) Axis induction by β-Catenin mRNA (80 pg) in Xenopus was blocked by co-injection of HRS-MO, and partially rescued by 10 pg human HRS mRNA.
(P-R) Vps4-EQ, but not Vps4-WT, mRNA inhibited secondary axis formation by β-Catenin mRNA.
(S-X) Nuclear function of β-Catenin in Xenopus embryos. Wild-type, but not β-Catenin depleted embryos, contain neural tissue marked by Sox2 (n=44 and n=25). Microinjection of DN-GSK3 mRNA (150 pg, 4 times at 4-cell) dorsalized in a β-Catenin dependent manner (n=38 and n=28). DN-Tcf3 mRNA (200 pg, 4 times) blocked dorsalization by DN-GSK3 (n=36), while the fusion construct β-Catenin-DN-xTcf3 (30 pg) rescued the ventralizing effects of β-Catenin MO (n=26). Arrowheads indicate position of the blastopore.
(Y-Aa) Epistatic experiment showing that β-Catenin-DN-xTcf3 fusion protein does not require MVB formation to induce secondary axes in Xenopus embryos.
(Ba-Da) Epistatic experiment showing that the downstream target of Wnt signaling Siamois is not affected by MVB inhibition. |
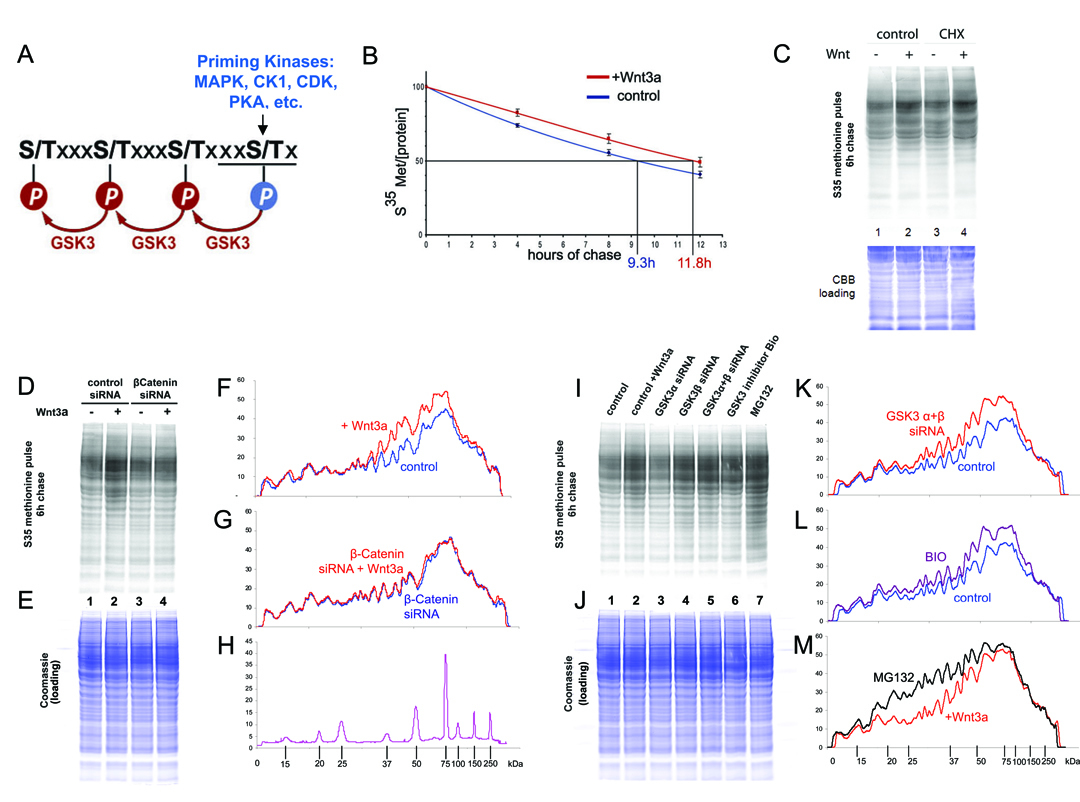 |
Figure 5. Wnt Prolongs the Half-Life of Cellular Proteins via GSK3 Inhibition
(A) 20% of human proteins contain three or more putative GSK3 sites primed by various kinases.
(B) Radioactive pulse-chase experiment showing that the half-life of total proteins was increased by Wnt3a treatment of 293T cells.
(C) Wnt3a treatment stabilized many proteins after 6 hours of chase in autoradiographs, even after inhibiting protein synthesis with Cycloheximide (see Figure S6A); Coomassie Brilliant Blue (CBB) staining provides equal loading control.
(D-H) β-Catenin was required for protein stabilization by Wnt3a. Densitometric tracings show that proteins stabilized by Wnt are within the 35 to 150 kDa range (note m.w. standards).
(I-M) Depletion of GSK3α/β or their chemical inhibition by BIO resulted in protein stabilization similar to Wnt3a, while proteasomal inhibition by MG132 stabilized a wider range of proteins than Wnt3a or GSK3 inhibition. |
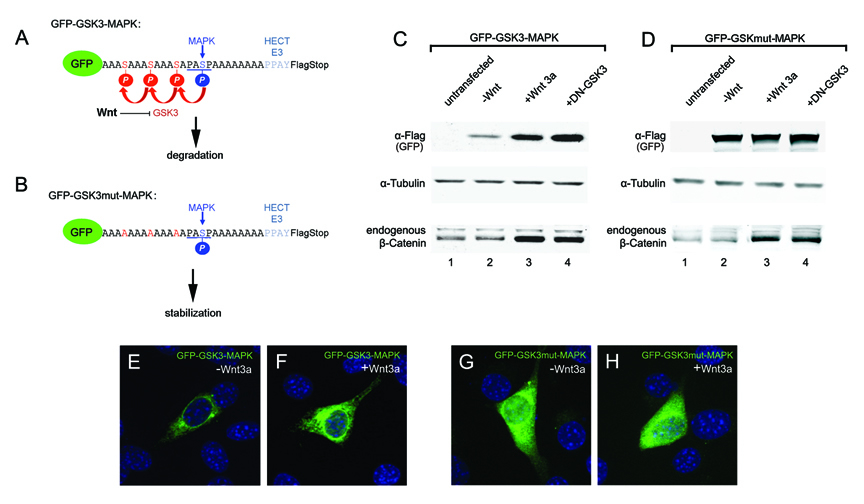 |
Figure 6. A Wnt-Regulated Reporter Protein Generated by Introducing Multiple GSK3 Phosphorylation Site
(A and B) Sequences of the GFP-GSK3-MAPK biosensor construct and its GSK3 phosphorylation-resistant mutant. This protein is predicted to be primed by MAPK/Erk and further phosphorylated by GSK3.
(C) Western blot showing stabilization of GFP-GSK3-MAPK and endogenous β-Catenin by Wnt3a treatment (6 hours) or DN-GSK3 transfection in transfected 3T3 cells. α-Tubulin served as loading control.
(D) When GSK3 sites were mutated, GFP was stabilized and did not respond to Wnt.
(E-H) GFP fluorescence in 3T3 cells transfected with GFP reporter proteins. Note that fluorescence was increased by Wnt3a treatment or mutation of GSK3 sites. |
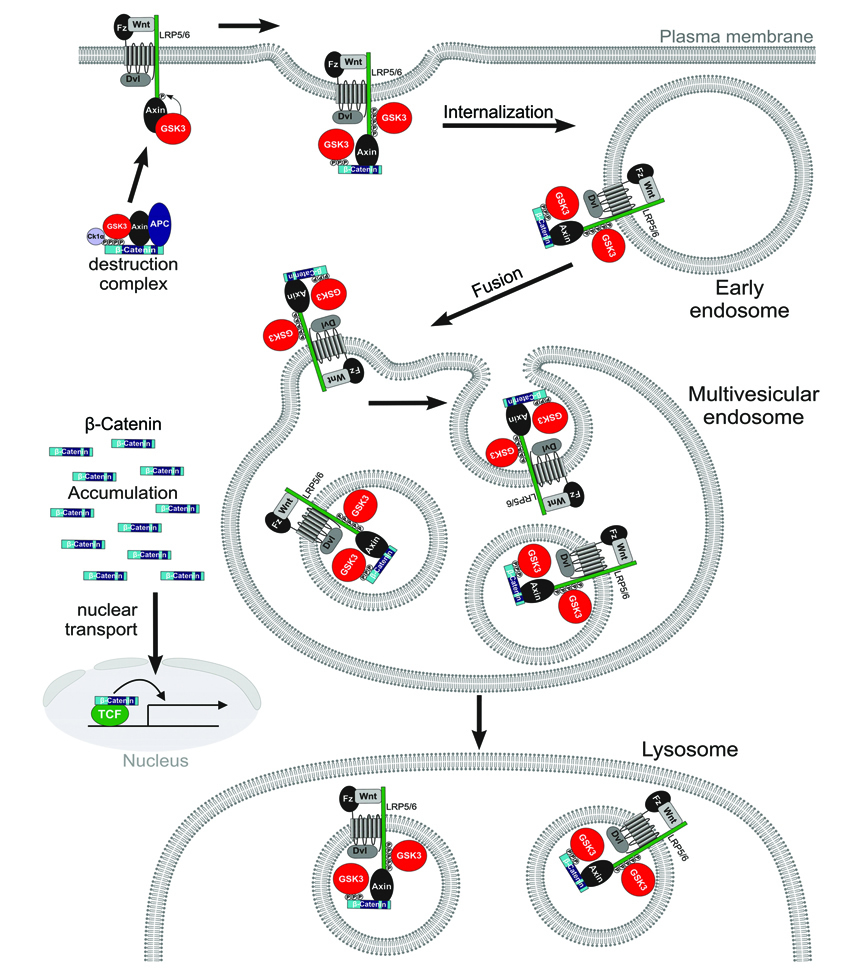 |
Figure 7. Model of Canonical Wnt Signaling through the Sequestration of GSK3 inside Multivesicular Endosomes
Binding of GSK3 (in red) to the Wnt receptor complex (including phospho-LRP6, phospho-β-Catenin, and other GSK3 substrates such as Dvl, Axin and APC) sequesters GSK3 inside small intraluminal MVB vesicles, causing its cytosolic substrates such as β-Catenin (in blue) and many other proteins to become stabilized (see text). The initial GSK3 molecules are recruited to the receptor complex bound to Axin, ensuring that the GSK3 fraction bound to the destruction complex is depleted first. Diagram modified from Zeng et al. (2008). |
________________________________________________________________________________________________________________________________________________________________________________
MITF drives endolysosomal biogenesis and potentiates Wnt signaling in melanoma cells
Diego Ploper, Vincent F. Taelman, Lidia Robert, Brian S. Perez, Björn Titz, Hsia-Wang Chen, Thomas G. Graeer, Erika von Euw, Antoni Ribas and Edward M. De Robertis
Proc. Natl. Acad. Sci. USA 112, E420-E429 (2015)
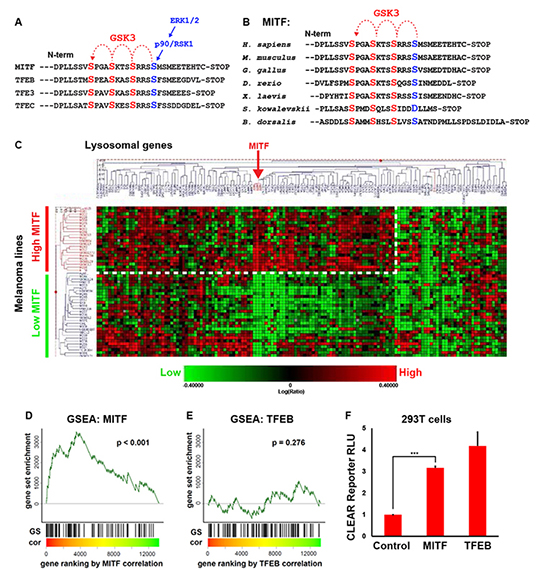 |
Fig. 1. MITF mRNA correlates with lysosomal gene expression in melanoma cell lines.
(A) MITF and the lysosomal master gene regulator TFEB have three putative C-terminal GSK3 phosphorylation sites, with a previously validated priming site.
(B) The C-terminal GSK3 sites on MITF have been highly conserved throughout evolution, including in the oriental fruit fly B. dorsalis.
(C) Heat map obtained from a RNA microarray panel of 51 melanoma cell lines, which cluster into 2 distinct groups, one with high MITF and the other with low MITF expression, when queried for a panel of 89 lysosomal genes. The group with high MITF expression (which includes all cell lines with MITF genomic amplifications) upregulates many, but not all, lysosomal genes (dashed line).
(D and E) Gene Set Enrichment Analysis (GSEA) of an expression data set consisting of a panel of 83 additional, different, melanoma lines confirms that MITF, but not TFEB, significantly correlates with the lysosomal gene set in melanoma. Microarray data for melanoma cell lines was obtained from Hoek et al. (30). Genes were ranked by their correlation (cor) with MITF or TFEB (red to green = high to low correlation). The positions of lyosomal genes (Sardiello et al., 2009) among over 12,000 genes per cell line are marked as vertical lines (GS). Enrichment of the lysosomal gene set at the top of the ranked lists was assessed with a permutation based Kolmogorov-Smirnoff (KS) non-parametric rank test. A significant correlation for MITF, but not for TFEB, was found.
(F) Transfection of MITF activated a CLEAR element-Firefly luciferase reporter (22), in transient transfections of HEK 293T cells. Renilla luciferase driven by the CMV promoter was used for normalization purposes.
|
 |
(A) Schematic diagram of the C32 Tet-inducible MITF melanoma cell line (20).
(B) Strong induction of MITF after Tet treatment was detected in C32 cells by western blot with anti-MITF antibody.
(C-D’) Increase in the number of vesicular structures upon MITF induction observed by DIC light microscopy. Note vesicular structures seen at high power (arrows).
(E and F) MITF induction increases immunostaining of the late endosomal marker LAMP1.
(G) Quantification by flow cytometry of the increase in LAMP1 levels upon MITF induction.
(H and I) MITF induction increases immunostaining of the MVB marker CD63.
(J) Quantification by flow cytometry of the increase in CD63 upon MITF induction. (K) MITF induction increased the transcripts of many lysosomal genes containing CLEAR elements in C32 melanoma cells, as validated by RT-qPCR. MITF induction in C32 melanoma cells upregulated transcripts for the CLEAR element lysosomal genes a-N-acetylglucosaminidase (NAGLU), chloride channel voltage sensitive 7 (CLCN7), prosaposin (PSAP), cathepsin D (CTSD), cathepsin A (CTSA), sialidase 1 (NEU1), a-galactosidase (GLA), mucolipin 1 (MCOLN1), b-glucocerebrosidase (GBA), and serine carboxypeptidase 1 (SCPEP1). Error bars indicate the standard error of the mean from three independent experiments.
|
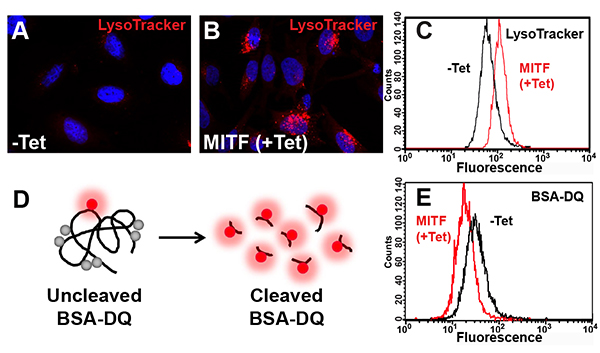 |
(A and B) Acidic organelles visualized by treatment of living cells with LysoTracker dye.
(C) Quantification by flow cytometry of the increase in acidic organelles observed upon MITF induction.
(D) Schematic representation of the BSA-DQ reagent used for detecting lysosomal proteolytic activity. BSA-DQ added to the culture medium is endocytosed, but only fluoresces when cleaved by proteases inside lysosomes.
(E) MITF induction decreases lysosomal activity, as quantified by flow cytometry.
|
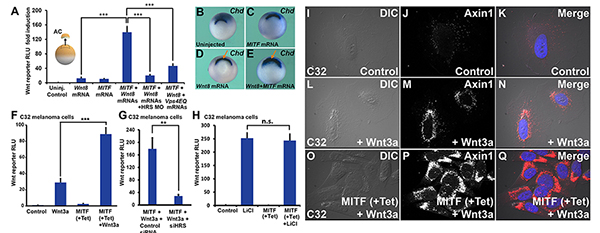 |
(A) MITF enhances Wnt signaling in Xenopus ectodermal explants. This enhancement is ESCRT-dependent, as it is blocked by HRS/Vps27 morpholino, or a dominant-negative form of the Vps4 ATPase (HRS MO and VPS4EQ) (*** p < 0.001).
(B-E) MITF cooperates with a low dose of Wnt8 mRNA, expanding the Spemman organizer (arrows), the region that expresses chordin mRNA in Xenopus whole-mount in-situ hybridization.
(F) MITF induction increases Wnt signaling in the C32 MITF-inducible melanoma cell line. The Wnt BAR Firefly luciferase reporter and EF1a-driven Renilla luciferase by were permanently introduced with lentivectors into the C32 cell line. (G) HRS/Vps27 knockdown by HRS siRNA decreased Wnt signaling in Tet-induced C32 cells (** p < 0.01).
(H) MITF induction did not affect Lithium chloride-induced b-Catenin signaling.
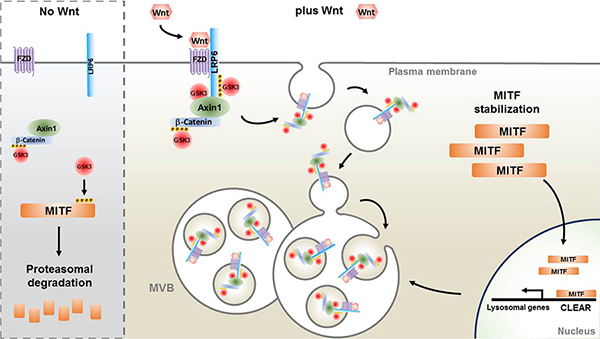
(I-Q) Immunostaining for Axin1, the key scaffold of the b-Catenin destruction complex. Note that Wnt signaling relocalizes Axin1 to vesicular structures, and that this effect is strongly enhanced by MITF induction with Tet. For relocalization of other Wnt components (phospho-LRP6, GSK3, and phospho-b-Catenin) after MITF induction and Wnt3a protein treatment, see Figs. S6 to S8. |
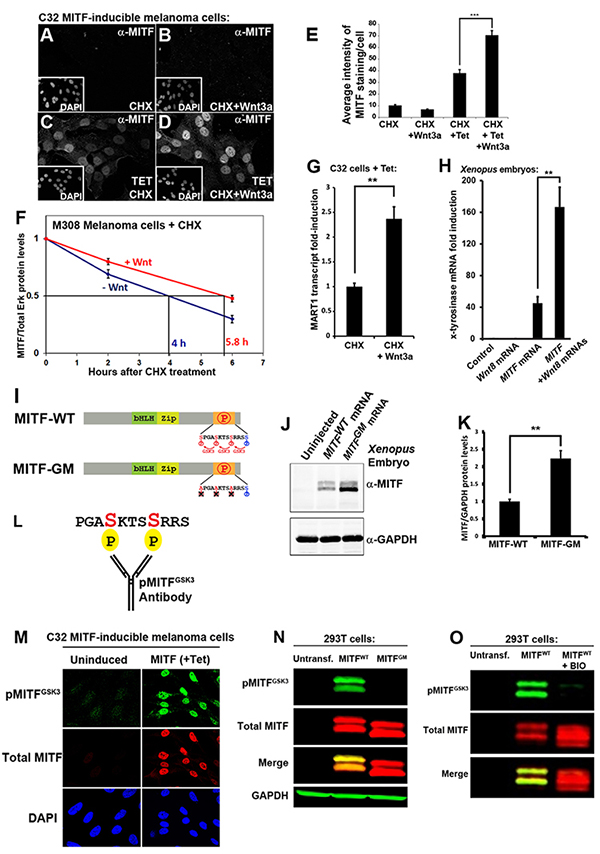 |
(A-D) MITF immunostainings in C32 cells. Wnt3a protein treatment for 5 hours stabilized MITF protein in the presence of cycloheximide (CHX).
(E) Quantification of MITF staining per nucleus from previous experiment (*** p < 0.001). (F) MITF protein levels (normalized to total Erk1/2) from 3 independent western blot experiments upon treatment of M308 melanoma cells with Wnt3a; Wnt prolongs the half-life of endogenous MITF protein. (G) RT-qPCR of the MITF target gene MART-1 in Tet-induced C32 cells treated with CHX and Wnt3a (** p < 0.01). (H) RT-qPCR for the MITF target gene x-tyrosinase obtained from Xenopus laevis embryos microinjected with Wnt8, MITF, or MITF + Wnt8 mRNAs. Wnt8 markedly increased MITF activity.
(I) Diagram depicting MITF wild type (MITF-WT) and a MITF GSK3 phosphorylation-resistant mutant (MITF-GM).
(J) Western blot of Xenopus laevis embryos injected with equal amounts of mRNA for MITF-WT or MITF-GM and blotted for MITF. GAPDH was used as a loading control.
(K) Quantification of western blots from three independent Xenopus experiments showing that MITF-GM is more stable than MITF-WT.
(L) Diagram of a pMITFGSK3 antibody raised against the indicated peptide corresponding to the C-terminal region of MITF with two phosphorylations.
(M) pMITFGSK3 antiserum mirrors the total MITF immunostaining pattern detected with an anti-MITF mAb in Tet-induced C32 cells. This indicates that the phospho-antiserum is specific for MITF.
(N) Western blot of HEK 293T cells transiently transfected with MITF-WT or MITF-GM and blotted for pMITFGSK3, total MITF, and GAPDH as a loading control. Note that MITF-GM is not recognized by the phospho-specific MITF antibody.
(O) Western blot of HEK 293T cells transiently transfected with MITF-WT treated with or without BIO, a specific GSK3 inhibitor. Note that pMITFGSK3, but not total MITF (anti-MITF mAb), is inhibited by BIO. |
 |
|
________________________________________________________________________________________________________________________________________________________________________________
Endocytic control of growth factor signalling: multivesicular bodies as signalling organellles
Radek Dobrowolski and Edward M. De Robertis
Nature Reviews Molecular Cell Biology 13, 53-60 (2012)
Signal transduction and endocytosis are intertwined processes. The internalization of ligand-activated receptors by endocytosis serves to attenuate signals by targeting receptors for degradation in lysosomes, but can also maintain signals in early signalling endosomes. In both cases, localization to multivesicular endosomes en route to lysosomes is thought to terminate signalling. However, during Wnt signal transduction, sequestration of the enzyme Glycogen Synthase Kinase 3 inside multivesicular endosomes results in the stabilization of many cytosolic proteins. Thus, the role of endocytosis during signal transduction may be more diverse than anticipated, and multivesicular endosomes may constitute a crucial signalling organelle.
|
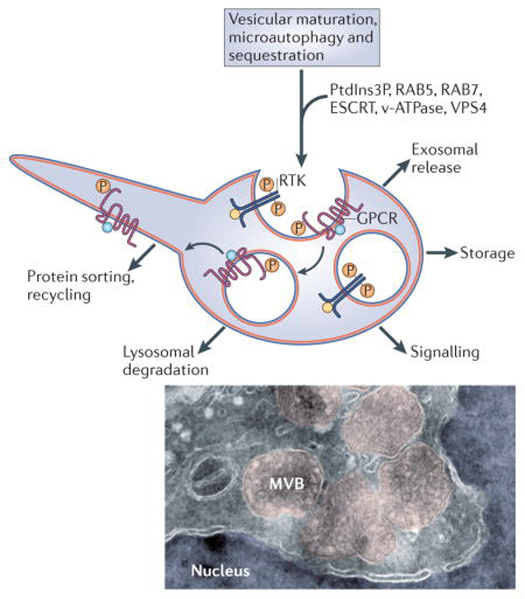
Box 1 │Diverse functions of multivesicular endosomes
Multivesicular endosomes are characterized by the internalization of small intraluminal vesicles (ILVs) of about 50 nm. This requires the orderly recruitment of components of the ESCRT machinery4,70. In addition, ILV formation requires the endosome-specific lipid phosphatidylinositol-3-phosphate (PI3P), and the Vps4 AAA-ATPase to pinch-off the vesicles71,72. The matrix of endosomes is gradually acidified by v-ATPses as they undergo maturation, enlarge and convert the early Rab5 compartment into Rab7 late endosomes73,74. The lumen of early recycling endosomes has a pH of 6.5 -6.4 (compared to 7.2 in cytosol), late multivesicular endosomes a pH of 6.0-5.0, and after fusing with lysosomes pH of 5.0-4.5 is reached75,76. Lysosomal hydrolases degrade proteins and lipids at acid pH. The diverse functions of multivesicular endosomes are indicated. In addition to serving as precursors for lysosomal degradation77,78, ILVs can be released into the extracellular space as exosomes when the entire organelle fuses to the plasma membrane79-81. The sequestered proteins can also be transiently stored and recycled back to the cytoplasm or the plasma membrane via back-fusion of ILVs to the peripheral endosomal membrane82 and membrane recycling through tubular structures26. Membrane proteins are sorted into ILVs after becoming mono-ubiquitinated6. Cytosolic material can be engulfed into multivesicular endosomes by microautophagy, which involves invagination of larger vesicles containing cytoplasmic components such as ribosomes83. The electron micrograph illustrates the morphology of multivesicular bodies (shadowed in pink). These MVBs were induced by a constitutively-active form of LRP6 receptor that generates a very strong Wnt signal by sequestering GSK3 inside these structures12. Cryo electron microscopy image courtesy of David D. Sabatini, New York University. |
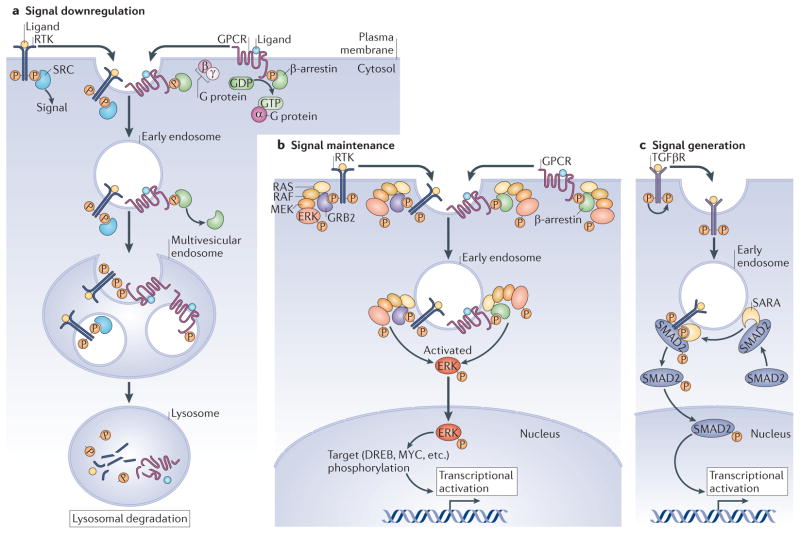 |
Figure 1│ Current models for the intersection between endocytosis and signaling pathways.
Three major models for the crosstalk between endocytosis and cell signaling: a│ Downregulation of signals though degradation of receptor complexes. RTKs and GPCRs signal at the plasma membrane through their effectors or activated G-proteins, which are released as early endosomes acidify. After trafficking of receptors into the intraluminal vesicles (ILVs) of multivesicular endosomes, these organelles fuse with lysosomes and receptors are degraded. b│ MAPK signaling provides an example in which signaling is maintained by early endosomes. The signal is generated at the plasma membrane and continues signaling in early endosomes through formation of the Ras-Raf-Mek-Erk complexes bound to the Grb2 adaptor for RTKs or β-Arrestin for GPCRs. Activated Erk is released from the endosome and translocates to the nucleus to phosphorylate its targets. c│ For TgfβR, as well as other receptors, the signal is generated on signaling endosomes. Sara is a FYVE domain (PI3P-binding) protein that recognizes the activated receptor and recruits the Smad2 transcription factor to signaling endosomes. Phospho-Smad2 is then released, entering the nucleus to promote transcriptional activation. |
|
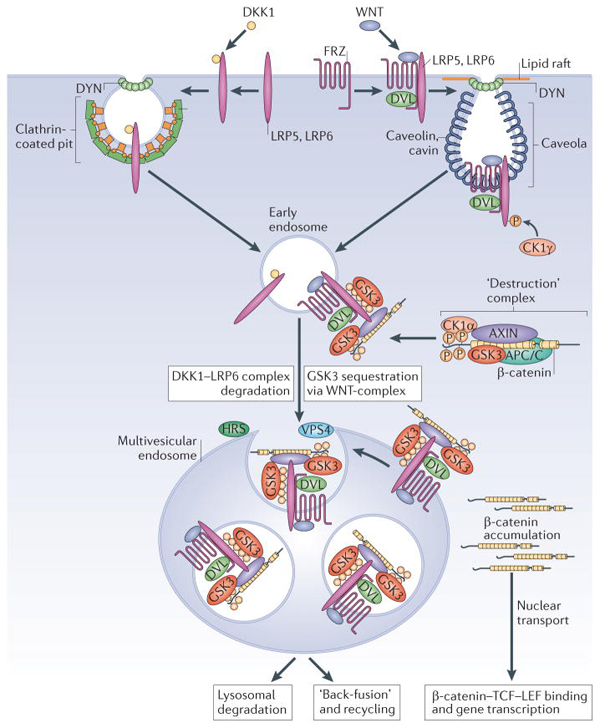
Figure 2 │ Endocytosis is required for canonical Wnt signaling.
Two Wnt co-receptors, Frizzled (Frz) and LRP6, are required in the plasma membrane for canonical Wnt signaling. When cells are exposed to the antagonist Dkk1, LRP6 is directed to endocytosis by clathrin-coated vesicles, becoming inactive and subsequently degraded27. Conversely, when the cell receives a Wnt signal, Wnt binds to both LRP6 and Frz. This recruits Dishevelled (Dvl), which is required for polymerization of the co-receptors in lipid rafts and endocytosis through caveolin-containing vesicles13. A requirement for the endocytosis effector Dynamin (Dyn) and v-ATPase have also been demonstrated21,84. The cytoplasmic tail of LRP6 is phosphorylated by CK1γ and GSK3, activating endocytosis85 and binding components of the β-Catenin destruction complex (Axin, GSK3). Early endosomal vesicles mature into multivesicular endosomes (in a process that requires HRS and Vps4) and Wnt receptor complexes are internalized, sequestering GSK3 (in red) inside ILVs. Sequestration of the GSK3 enzyme in multivesicular endosomes is an essential step required for sustained Wnt signaling. The decrease in active GSK3 causes newly-synthesized β-Catenin to accumulate, which is transported to the nucleus where it activates gene transcription by Tcf/LEF. |
|
Figure 3│ Generation of protein half-life biosensors and their potential use in signaling integration studies.
a│A generic biosensor of intracellular GSK3 activity consisting of a fluorescent protein (GFP or RFP) with a C-terminal peptide containing phosphorylation sites. Three artificial GSK3 sites (in red) were placed in front of a priming kinase site (which could correspond to MAPK, CK1, CK2, PKA or many others); further downstream an epitope-Tag (useful for biochemical analyses) and a stop codon complete the biosensor. The addition of GSK3 phosphorylation sites to the protein destabilizes it, forming a short phosphodegron that promotes polyubiquitinylation by endogenous E3 ligases and its proteasomal degradation.
b│ Phosphorylation sites that promote degradation in human β-Catenin protein and in the artificial GFP biosensor, GFP-GSK3-CK1, derived from its sequence12. CK1 primes three phosphorylations by GSK3. |
|
Figure 4│ Hypothesis: the dorsal determinants of the Xenopus eggare endosomal components.
The vegetal pole of the frog egg contains “maternal determinants” of unknown composition. We propose here that they may correspond to Wnt-containing early endosomes that become incorporated into multivesicular bodies on the dorsal side of the zygote. This would sequester GSK3 and Axin inside MVBs, triggering the earliest Wnt signal in vertebrate embryonic differentiation, which induces the Nieuwkoop signaling center. In the unfertilized egg, membrane vesicles are observed in the vegetal pole61. The sperm brings with it the centriole, which organizes a cortical network of microtubules. Early endosomes from the vegetal pole are transported along microtubules to the dorsal side, which forms opposite the sperm entry point. The new axis forms where vegetal material and animal cytoplasm mix55. Cortical microtubules cause a rotation of the egg cortex towards the sperm entry point, displacing the superficial pigment of the egg53. This forms a lighter “dorsal crescent” that marks the dorsal side. Arguments supporting the sequestration of GSK3 in multivesicular endosomes as the cause of the earliest Wnt signal are discussed in the main text. |